
Nous avons donc vu la semaine dernière le modèle évolutif, conçu par Richard Prum, qui tente de retracer l'histoire des différentes étapes évolutives menant à l'émergence de plumes adaptées au vol. Ce modèle, si vous n'avez pas la patience de lire mon précédent article (mais c'est très mal!), le voici:

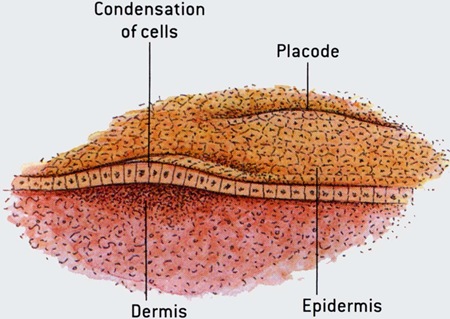
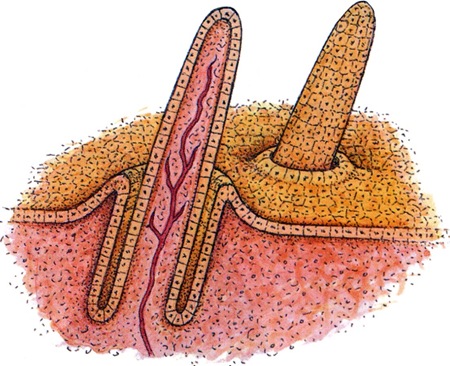
Dans ce modèle, cinq étapes se succèdent et sont récapitulées lors du développement d'une plume chez un oiseau. En effet, chez un poulet par exemple, toutes les plumes sont issues d'une placode puis d'une plume folliculaire qui évoquerait peut être la morphologie des toute premières plumes (les proto-plumes). Les barbes et barbules apparaissent également de manière séquentielles. Enfin, en fonction de la manière dont les barbes croissent, parallèlement ou de manière hélicoïdale, la plume prendra une forme particulière, duveteuse ou penne respectivement.
Tout ceci suggère que chacune de ces structures sont issues d'innovations évolutives successives. En effet, pour qu'une plume puisse se former, il faut nécessairement qu'on obtienne au début un tube. Les barbes des plumes duveteuses ou pennes requièrent donc l'existence préalable d'une structure de type plume folliculaire. de la même manière, le rachis d'une plume penne provient de la fusion des barbes qui croissent de manière hélicoïdale: le rachis nécessite donc l'innovation évolutive des barbes puis celle de la croissance hélicoïdale. Vous voyez ou je veux en venir: chaque étape du modèle n'a pas été postulée au hasard! Chacune équivaut à l'ajout d'une innovation évolutive. Et la grande élégance de ce modèle vient du fait que chaque étape correspond à une prédiction testable!
De nos jours, de simples données développementales ne sont plus satisfaisantes pour étayer un modèle évolutif. Enquêter sur l'évolution des plumes requiert la description des mécanismes moléculaires à l'origine de ces étapes développementales. J'appuie sur l'expression à l'origine parce qu'elle reflète bien la démarche de l'évo-devo: découvrir les protagonistes moléculaires qui contrôlent des étapes du développement revient à découvrir les outils ancestraux qui ont servi à mettre en place les premières esquisses de structures aussi complexes qu'un plume.
C'est chaud à piger, hein? Allez, vous inquiétez pas... c'est juste le début!
Pour comprendre un petit peu la démarche qu'ont entrepris les chercheurs qui voulaient étudier l'histoire évolutive des plumes, il faut d'abord savoir ce que sont exactement les oiseaux. Par exemple, savez vous quels sont les animaux actuels qui sont les plus proches des oiseaux en terme de parenté? Et bien de nombreuses études morphologiques et moléculaires ont permis de déterminer qu'il s'agissait des crocodiles! Crocodiles et oiseaux forment donc un groupe qu'on appelle les archosauriens.

Etonnant, non?
C'est donc en comparant les crocodiles et les oiseaux qu'on peut commencer à accumuler des indices sur l'origine des plumes. Il faut donc chercher une structure cutanée partagée par les crocodiles et les oiseaux qui pourraient avoir conservé des propriétés ancestrales chez l'un ou l'autre groupe d'espèces: ça ne vous dit rien? Et si je compare ça par exemple:


Est-ce que l'écaille conservée chez ces archosaures pourraient être la structure ancestrale qui aurait évolué en plume dans la lignée des oiseaux?
Pour en avoir le cœur net, il faut comparer le développement de ces structures et surtout les gènes impliqués dans leur formation. En effet, c'est en menant une analyse comparative du développement des écailles d'oiseaux et de crocodiles que l'on peut commencer à comprendre quels ont pu être les innovations développementales qui ont permis leur complexification lors de l'histoire évolutive des plumes.
C'est pour la première fois je crois que je vais évoquer sur SSAFT le nom de certains gènes et il faut que je vous prépare psychologiquement à la chose: rappelez-vous les biologistes sont de grands enfants (si vous avez des doutes en voici les confirmations ici et là ).
Il s'avère que chez la plupart des vertébrés, deux gènes interviennent très souvent pour la formation des structures cutanées comme les poils, les écailles et les plumes. Ces deux gènes sont Shh (pour Sonic Hedgehog, oui oui, le personnage de SEGA) et Bmp2 (pour Bone Morphogenetic Protein 2... bon ben ça arrive que les biologistes soient sérieux c'est tout!). Il a fallu donc déterminer comment agissent ces gènes lors de la formation des écailles des archosaures.
Vous en avez peut être déjà entendu parlé, mais par prudence, je réexplique: bien que toutes les cellules d'un organisme partage les même gènes, elles sont différentes les unes des autres parce qu'elles n'activent pas leurs gènes de la même manière. Ainsi, au sein d'un groupe de cellules identiques, on peut obtenir une très grande variété de cellules finales avec des programmes d'activation de gènes différents. En génétique, quand un gène est activé on dit qu'il s'exprime. L'expression d'un gène peut être suivie à l'échelle d'une cellule, d'un organe voire même d'un organisme entier à l'aide d'une technique qu'on appelle l'hybridation in situ et qui permet de colorer (le plus souvent en bleu) des cellules exprimant un gène spécifique.
Etudions donc, si vous le voulez bien, l'expression des gènes Shh et Bmp2 lors de la formation des écailles d'oiseaux (poulet et canard) et de crocodiles, mais aussi lors de la formation des placodes des plumes de ces volatiles:

En ce qui concerne les écailles de tous ces archosaures, l'expression de ces deux gènes est presque identique: Shh se retrouve exprimé dans les cellules du bas de l'écaille en formation tandis que Bmp2 s'exprime dans le haut de l'écaille. C'est d'ailleurs ce qui est schématisé à droite avec les rectangles où le côté haut est bleu (Bmp2) et le côté bas est rouge (Shh). Ces deux expressions sont même assez similaires à l'expression de Shh et Bmp2 dans la plume au stade placode chez le poulet et le canard. Que suggèrent donc ces résultats?
Et bien il est probable que Shh et Bmp2 étaient employés de la même manière dans les écailles du dernier ancêtre commun des oiseaux et des crocodiles, et que cette manière de s'exprimer a été maintenue dans le stade placode lors de la formation des plumes.
Ces première données moléculaires nous confirment donc que les plumes proviennent probablement d'écailles chez un ancêtre (la morphologie de ces écailles n'ayant que peu changé chez les crocodiles). La première étape de l'histoire évolutive des plumes est récapitulée par l'étape développementale du stade placode.
Continuons notre inspection: comment s'expriment nos nouveaux gènes préférés dans les étapes suivantes de la formation des plumes?

Expression des gènes Shh et Bmp2 lors de la morphogenèse de plumes duveteuses (D-E) et pennes (F-H)
Les images A et B nous montrent que Shh (A) et Bmp2 (B) sont exprimés dans un territoire diffus chevauchant au stade plume folliculaire. Tiens, tiens, c'est différent de ce qui se passait au stade placode précédent où les territoires d'expression étaient bien distincts. Et puis l'image C nous montrent que cette expression (ici celle de Shh) change au fur et à mesure que la plume mature: on voit apparaitre des stries. C'est curieux parce que des stries, c'est ce qui pourrait préfigurer l'émergence de barbes! Si c'est le cas, on devrait voir une différence de ces stries pour des plumes duveteuses et des plumes pennes: des stries bien parallèles pour une plume duveteuse (D) et en spirale pour des plumes pennes avec un côté ou les stries se rejoignent (F) et une arrête sans stries de l'autre côté (G). Et ben voilà, bingo: on a deux gènes qui peuvent nous permettre d'observer la préfiguration des barbes au cours de la formation des plumes.
Mais pour en être certain, Matthew Harris, John Fallon et Richard Prum ont voulu soumettre cette hypothèse à un test : est-ce que l'expression de ces deux gènes peut expliquer la variété de plumes qui parcourent le corps des jeunes poussins à la naissance. En effet, le duvet des poussins est souvent très divers avec des plumes bifurquées, trifurquées, fusionnées, ou même complètement séparées de la peau du poussin. Les auteurs ont donc étudié attentivement l'expression du gène Shh sur de très nombreux follicules et ont obtenu ces images saisissantes (gardez en tête que l'expression de Shh marque la limite des barbes et que celles-ci paraissent donc blanches à l'image):
 L'expression de Shh et Bmp2 préfigure les variations
L'expression de Shh et Bmp2 préfigure les variations
morphologiques des plumes du duvet de poussin
Chaque petites anomalies dans la régularité des stries explique une morphologie particulière de plumes: l'image C par exemple montre une bifurcation qui laisse apparaitre une barbe non rattachée à la base du follicule et le résultat en H montre une mini plume qui se détache du reste du duvet...
Ce qui est quand même très curieux, c'est que ce soit les même deux gènes, Shh et Bmp2, qui sont impliqués dans ces différentes étapes du développement de la plume. En tout cas, un petit schéma s'impose pour résumer tout ça:

L'emploi répété de la combinaison de gènes Shh et Bmp2 n'est finalement pas si curieuse que ça. En effet, ces deux gènes constituent ce qu'on appelle un module génétique, une relation entre deux gènes conservée au cours de l'évolution mais pouvant être employée pour générer des modifications morphologiques significatives.
En l'occurrence, les chercheurs ayant étudié l'expression des gènes Shh et Bmp2 lors de la morphogenèse des plumes pensent que ce module génétique a été utilisé plusieurs fois au cours de leur évolution et corroborent le modèle de Richard Prum jusqu'à l'étape IIIa.
Au départ, l'ancêtre des archosaures employait ce module lors de la formation d'écailles. Des plumes tubulaires primitives ont évolué à partir de ces écailles par le recrutement du module génétique Shh-Bmp2 dans un domaine chevauchant d'un côté de l'écaille. L'étape suivante implique un nouveau recrutement de ce module pour définir des domaines d'expression longitudinaux qui strient la plume folliculaire: résultat, on obtient la toute première plume à barbes, une plume duveteuse. Enfin, une variation du module entrainant des stries non plus longitudinales mais hélicoïdales permet l'émergence d'une plume penne possédant un axe, le rachis.
Bien entendu, avec de nombreuses expériences dont je vous passerai la description, les auteurs ont montré que Shh et Bmp2 étaient essentiels à chacune de ces étapes et que la perturbation de leur activité entrainait une perturbation spécifique de la morphologie de la plume selon l'étape développementale.
Ce qui manquait au tableau, c'était le type de relation qu'entretiennent ces gènes au sein du module. Encore une fois, un modèle a été conçu avant d'être testé empiriquement. Bon autant vous prévenir tout de suite, ce modèle est à peine compréhensible. Le postulat de départ est le suivant: pour créer des stries d'expression, il est possible d'employer un simple module impliquant deux gènes dont un jouerait le rôle d'activateur (Shh qui promouvrait sa propre expression et l'expression de son inhibiteur) et l'autre le rôle d'inhibiteur (Bmp2 qui cherche à diminuer l'expression de Shh). En réalisant des simulations à partir de différentes conditions de départ, voici les résultats obtenus:
 Le modèle inhibiteur-activateur est suffisant pour expliquer
Le modèle inhibiteur-activateur est suffisant pour expliquer
l'émergence des stries d'expression et donc de la formation de barbes
Sachant que Shh et Bmp2 portent bien ces fonctions, ce modèle a donc été confirmé et renforce le modèle évolutif des plumes employant à plusieurs reprises ce module génétique. Un dernier obstacle restait à franchir: celui expliquant les stries hélicoïdales (images I et J ) qui permettent la formation de plumes pennes. Les auteurs ont estimé qu'un troisième protagoniste était nécessaire pour expliquer ce type d'expression, favorisant d'un côté les bifurcations (I) et de l'autre l'arrêt des stries (J).
Il s'avère que quelques mois après avoir publiés leur modèles, un groupe de chercheurs trouvaient de manière totalement indépendante ce troisième protagoniste, le gène Wnt3a.
Bref, ça y'est j'ai perdu les 90% d'entre vous à partir de maintenant... C'est pas grave je compte vous récupérer la semaine prochaine! Comment? Et bien tout simplement en vous montrant comment les données paléontologiques corroborent soutes ces données développementales et moléculaires...
C'est bien ça: la semaine prochaine, c'est Jurassic-à-plumes Park!
Références :
Harris, M. P., J. F. Fallon, et al. (2002). "Shh-Bmp2 signaling module and the evolutionary origin and diversification of feathers." J Exp Zool 294(2): 160-76.
Harris, M. P., S. Williamson, et al. (2005). "Molecular evidence for an activator-inhibitor mechanism in development of embryonic feather branching." Proc Natl Acad Sci U S A 102(33): 11734-9.
Prum, R. O. (1999). "Development and evolutionary origin of feathers." J Exp Zool 285(4): 291-306.
Prum, R. O. (2005). "Evolution of the morphological innovations of feathers." J Exp Zoolog B Mol Dev Evol 304(6): 570-9.
Prum, R. O. and A. H. Brush (2002). "The evolutionary origin and diversification of feathers." Q Rev Biol 77(3): 261-95.
Prum, R. O. and J. Dyck (2003). "A hierarchical model of plumage: morphology, development, and evolution." J Exp Zoolog B Mol Dev Evol 298(1): 73-90.
Yue, Z., T.-X. Jiang, et al. (2006). "Wnt3a gradient converts radial to bilateral feather symmetry via topological arrangement of epithelia." Proceedings of the National Academy of Sciences of the United States of America 103(4): 951-955.









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1 De Tom Roud - 19/05/2009, 13:35
Je ne connais pas les détails du modèle de formation de motif, mais vu la description que tu en fais, c'est sans aucun doute un modèle très classique de réaction-diffusion type modèle de Turing. En fait, c'est assez agaçant de voir que dès qu'il y a un motif en biologie, les gens sortent un modèle de Turing, alors que dans pas mal de cas, cela ne peut pas marcher car il n'y a pas de diffusion par exemple. J'avais expliqué ce problème dans un billet sur la segmentation :
http://tomroud.com/2007/04/12/physi...
2 De taupo - 20/05/2009, 09:50
@Tom Roud :
Shh et Bmp2 sont bel et bien des protéines diffusibles. Le modèle en question est le modèle activateur-inhibiteur qui prend comme hypothèse que :
(i) the inhibitor down-regulates activator function
(ii) the activator up-regulates its own expression
(iii) the activator increases the activity of the inhibitor.
Ces relations fonctionelles ont été démontrées dans cet article.
3 De Daniel Jonas Parker - 20/05/2009, 14:42
Les gènes segmentaires vous yavait pensé?
Les archosaures sont apparu il y a 290 millions d'années. Les dinosaures du types Tyrannosaures ne sont peut-être pas de bon candidats pour symboliser les théropodes non aviens : 6m de haut , 13m de long , 6 à 7tonnes .
Mais bon ils partagent la même conception des cellules dermiques avec les ptérosauriens, branche qui s'est séparés d'eux voici 240 millions d'années. Partageant un même caractère commun si lointaint soit-il est la preuves d'existence de proto-plumes sur Tyrannosaurus Rex
4 De Tom Roud - 21/05/2009, 16:27
OK, mais ce que je déplore, c'est que dès qu'il y a formation de motif en biologie, les théoriciens viennent "prédire" que c'est un mécanisme de réaction-diffusion, ce qui est un peu énervant à la longue, d'autant plus quand cela s'avère faux au bout du compte.
Je me souviens d'une partie théorie d'un meeting d'evo-devo où tout le monde parlait constamment de réaction-diffusion, sans avoir identifié le moindre acteur moléculaire, alors que par exemple on sait que pour la segmentation, cela ne marche pas justement. Meinhardt, un des auteurs du papier que tu cites, est d'ailleurs bien placé pour savoir cela car il est LE chercheur qui a popularisé ces modèles en biologie, et en même temps, justement, son modèle de segmentation de la drosophile s'est avéré être faux ce qui lui a causé un peu de tort auprès de certains biologistes (malheureusement, car il est l'un des théoriciens qui connaît le mieux la biologie, et personnellement, je l'aime beaucoup).
5 De Daniel Jonas Parker - 09/06/2009, 11:49
Le sinosauptéryx est un bon candidat au titre du plus vieil acquéreur de proto-plumes pour un théropode non aviens , 250 millions d'années plus tard apparaisent les oiseaux dignes représentants du régne coelurosaurien.
les génes segmentaires sont de bons point de départ pour une étude approfondie du génome avien.
6 De taupo - 09/06/2009, 12:00
@Daniel Jonas Parker :Est-ce que vous pouvez un peu développer votre propos, svp? Pourquoi les gènes segmentaires, par exemple...
7 De Daniel Jonas Parker - 11/06/2009, 10:19
Lors du dévelopement de l'embryon les génes interférants ou génes segmentaires comme j'aime les appeler
8 De Daniel Jonas Parker - 11/06/2009, 10:27
L'ARN messager à dû être chambouler pour changer ansi la structure épidermique des archosaures , d'ou mon hypothèse : il ya 250 millions d'années les premiers dinosauriens primitif pointent le bout de leur museau , encodé dans leur patrimoine ADN le géne Shh subit d'importante modifs , l'info passe mal et l'ARN ne reconnaît pas la demande production de protéine altérée , dinos à plumes
9 De vélo - 12/06/2009, 11:30
@Daniel Jonas Parker : T con jonas je pense que le l'archéoptéryx mérite mieux ce titre car il est génétiquement placé(d'apres des recherches en mongolies et en argentine) dans l'aire du crétacée avec une apparition il ya 256 millions d'années.
10 De Daniel Jonas Parker - 15/06/2009, 15:55
Vélo je te rappelle que l'archéoptéryx est apparu il y a 150 millions d'années, or d'après plusieurs scientifiques cette animal est un cul de sac de l'évolution et je te rappelle qu'il y a256 millions d'années ce n'est pas le crétacé c'était le trias. Alors avant de dire des conneries pas possibles il vaudrait mieux que tu te renseignes sur l'histoire géologique de notre planète!!!
11 De Daniel Jonas Parker - 22/06/2009, 16:57
hn
12 De vélo - 24/06/2009, 13:24
créateur du site : Daniel Jonas Parker
13 De Daniel jonas parker - 24/06/2009, 14:54
je suis une merde.
14 De taupo - 24/06/2009, 23:42
@Daniel jonas parker : (alias velo déguisé...) Bon bon bon, je ne suis pas du genre à censurer les personnes désirant s'exprimer sur mon site, mais là, franchement, c'est tentant... principalement pour vous préserver de la honte que génère ce genre de dialogue stérile. Déjà, je comprends à peine de quoi il retourne dans vos commentaires (à Daniel Jonas Parker et Velo) tant vos efforts pour être compréhensibles sont minimes. Si en plus vous commencez à vous insulter gratuitement, et à usurper des identités... Les quelques minutes qui vous ont fallu pour faire ça auraient été bien mieux dépensées à réfléchir à un véritable argument à proposer l'un à l'autre, voir juste à un bout de phrase qui pourrait rendre intelligible votre différent aux autres lecteurs (moi compris) de ce blog.
Enfin moi j'dis ça, j'dis rien (c'est vrai ça, je dis pas grand chose sur le blog... pourquoi? Haha, bientôt une grande news les amis!)
15 De Daniel Jonas Parker - 25/06/2009, 15:06
Vous avez raison je suis une très très grosse merde de chien!!!!!!!!!!!!!!!!!!!!!!!
16 De Daniel Jonas Parker - 25/06/2009, 15:08
Mais je reste!
17 De vélo - 29/06/2009, 13:00
DJP=dijoncté=daniel jonas parker=.......
qui trouvera la réponse?
18 De vélo - 01/07/2009, 13:25
Je voudrais dire 2 mots au gars ayant usurpé mon identité je ne suis pas une merde et je voudrais rappelé que ce genre est fait pour l'échange de connaissance et non pour les insultes gratuite
19 De Daniel Jonas Parker - 06/07/2009, 09:09
Je sais! Mais je suis un gros con
20 De Jonathan Evans - 10/07/2009, 18:42
Hello, tout le monde je me prénome Jonathan j'ai était enchanté par l'article traitant sur l'implication direct de l'information génètique dans la formation des plumes
21 De jonathan evans - 13/10/2009, 11:52
@Jonathan Evans : gros con tu sait rien t nul
22 De s3ph|r0th - 15/10/2009, 12:51
Bon Kévin t'est gentil mais il y a des "skyblogs" pour exprimer ton mal de vivre, alors va jouer à la PSP et laisse nous entre hommes. Quelqu'un qui balance des insultes bien planqué derrière son écran, ça me rappelle mon chien qui aboie caché derrière le grillage mais dès qu'on s'approche il part en courant.
PS: à force d'être désagréable, tu t'est fait deux nouveaux ennemis, l'orthographe et la conjugaison! :p
23 De daniel jonas parker - 26/10/2009, 11:22
bonjour, je suis toujours moche et je n'ai pas d'amis et je slam sur les trotoirs
24 De s3ph|r0th - 30/10/2009, 10:46
En tous cas si l'évolution n'a pas dévellopé ton sens cognitif, elle t'a doté d'un remarquable sens de l'humour. Vérifie quand même si tu n'a pas des placodes, car vu ton étrange comportement tu est peut-être le successeur de l'homo sapiens sapiens. Et étant donné que JE l'ai repéré en premier, JE proposerais un nom pour cette espèce si la découverte se confirme.
25 De Daniel Jonas Parker - 05/11/2009, 10:17
je sui tré con et tré moche je vou emmerde tous gro conar
26 De daniel jonas parker - 10/11/2009, 11:47
@s3ph : bon je t emmerde je ne sui pa un homo sapien mais pluto un gro bonobo
27 De coeur d'encre - 04/01/2010, 11:29
rendez vous au bois de boulogne coté seine
28 De Velo - 06/11/2010, 17:11
Cher Daniel Jonas Parker tu devrais savoir que le bonobo est une espèce protégée et que l'on a pas le droit d'en posséder et encore moins de le faire se prostituer!!! N'as tu pas honte de te prostituer sur ce site visible par tous? Les gens vont êtres choqués!! cldt Velo.
29 De Velo - 06/11/2010, 17:11
Cher Daniel Jonas Parker tu devrais savoir que le bonobo est une espèce protégée et que l'on a pas le droit d'en posséder et encore moins de le faire se prostituer!!! N'as tu pas honte de te prostituer sur ce site visible par tous? Les gens vont êtres choqués!! cldt Velo.
30 De Daniel jonas parker - 16/11/2010, 14:20
Je ne suis pas un bonobos et au bois de boulogne y avait personne!!!!! Les con m ont elus roi des cons. Mes tarifs sont interessant!!!! VIVE LS SINGES
31 De Daniel Jonas Parker - 17/11/2010, 17:58
Bon que l'on arrete d'usurper mon identité !!! Je suis un gros bonobos sal mais c'est à moi de le dire !!!
32 De daniel jonas parker - 28/03/2011, 11:55
je vou emmerde tous
33 De vélocracheporte - 08/10/2011, 15:24
bonjour je suis un gros bonobo j'ai raté ma vie je suce des ***** et je chante comme une merde pc sous les yeux de mon pere j'ai appris a conduire et la moquette qui est sur ma tête est gratuite ( échantillon saint macloud)